
Neural correlates of psychedelic-induced delusions:
And other phenomenological symptoms of a dysregulated dopamine system

Introduction
In this text I will present the hypothesis (along with the facts and theories supporting it) that psychedelic induced delusions are caused by a runaway positive feedback loop between areas of the neocortex, the dorsal striatum and thalamus. I propose that this loop is driven by a hyperstimulation of dopaminergic receptors in the dorsal striatum as well as neuroplasticity in the neocortex (plasticity induced by 5HT2A stimulation). The loop would also be enabled due to disinhibition from hierarchically higher areas (likely the frontal lobe). This disinhibition occurs since the areas involved in top down control are overwhelmed by bottom up sensory information (also likely induced by 5HT2A stimulation). Furthermore, other personality changes present in some psychedelic users will be compared with dopamine driven psychopathology. The consequences of the hypothesis for psychedelic therapy and spontaneous spiritual awakening will be dwelt upon. Finally the ability of common neuroimaging techniques to verify the hypothesis will be reflected upon.
The dopaminergic neurochemistry of psychedelics
LSD is a direct agonist at D1 and D2 receptors.1 PET studies on pigs and subsequent in vitro studies show that LSD has a direct action at D2 and D3 receptors in pig striatum,2 Pharmacological studies on LSD-administration in rats in combination with various antagonists shows that LSD activates D4 receptors via a 5HT2A-receptor independent action 90 but not 30 minutes after LSD administration. Nichols connects the dots between this and a reported two-phasic phenomenology of the LSD-experience in man. First a psychedelic phase followed by a self centered and suspicious, sometimes paranoid phase, 4-6, sometimes up to 10 hours after drug intake,3 (remember that rats have a much faster drug-metabolism than humans and this can make translation of experimental results difficult). In human embryonic kidney cells modified to express the D4 receptor LSD acts as a full agonist.3
DMT has a stronger affinity to D1 than 5HT2A, however, its activity at D1 is presently unknown. It has no measurable affinity for D2 nor DAT4. To my knowledge direct agonism at dopaminergic receptors for psilocin has not yet been discovered, however, PET studies in rats strongly suggest that stimulation of 5HT2A (which all three drugs mentioned above are partial agonist at) indirectly leads to increased dopamine release in striatum5. DMT has been reported to increase the levels of the dopamine metabolite metabolite 3-methoxytyramine in humans during the experience but also a month after it. This may indicate an increased dopamine synthesis after repeated DMT use6. Since Ayahuasca always contains a MAO-inhibitor (most often harmaline) and MAO-inhibitors inhibits the enzyme which metabolizes monoamines (such as serotonin and dopamine) it goes without saying that this will increase the levels of dopamine in the synaptic cleft. Furthermore there’s tentative evidence that psilocybin induces dopaminergic activity in dorsal striatum from PET studies in human subjects7. Interestingly enough 5H2A and D2L (a splice variant of the D2 receptor) forms a hetero-receptor complex in striatum. This receptor complex is activated by LSD and DOI showing yet another way that psychedelics can promote dopaminergic signaling in striatum. 8,9 Other hetero-receptor complexes of relevance are the D1-D2 complex and the 5HT1A-D2 complex.10
Dopaminergic psychopathology and Parkinon’s disease
Dysregulated dopamine in striatum has been strongly correlated to attentional salience, delusional mood12, delusions of grandeur, paranoid delusions13,14, psychosis12, mania and hypomania15. A lot of the evidence for this comes from research into amphetamine induced psychosis, the mechanism of action of antipsychotics and various pharmacological research on people with bipolar depression.
Delusional mood is a mental state of perceived heightened awareness where everything feels like it has a hidden meaning and objects appear to be holy. There is a sense in the afflicted that he stands at the threshold of a great revelation, that he has the secrets of the universe at the tip of his tongue. In psychosis research this is considered to be a pre-psychotic stage, which can last for days or months before the great revelation is revealed, that would be the moment where the delusion crystalizes. Over time the delusion grows more and more rigid and unshakable, resistant to all logic and criticism12. Delusional mood is also a mystical state, which is pursued by many spiritual practitioners worldwide. Furthermore, it’s an intensely psychedelic state.
As mentioned above, The neural correlate of delusional mood is dopamine-overflow in striatum. What is the mechanism for this? A clue can be found in a mechanism for movement planning which has been revealed by research into Parkinson’s disease (PD). This research shows that the basal ganglia (a part of the striatum) works as a dopamine dependent filter for the selection of the correct movement16. When the decision to initiate a movement is made, premotor areas in the neocortex send suggestions of variety of movements to the basal ganglia where the correct movement, and the correct movement only, is filtered out and sent via the thalamus to motor areas (also in the neocortex) for further refinement and execution. The subthalamic nucleus is also involved in this circuitry. Feedback is then sent from motor areas to premotor areas closing the corticostriatal loop. When the dopaminergic axons which innervate the direct pathway in the basal ganglia from substantia nigra in the midbrain withers away in PD the result is muscle tremors and hypokinesia (slowed or inhibited movements), as the motor cortex is working from insufficient information (the correct motor signal can’t get through the noise so the net output from the basal ganglia onto the motor cortex becomes inhibitory. Too much dopamine (as in the peaks of levodopa fluctuations) can lead to dyskinesia (involuntary movements) also due to dysregulated dopaminergic balance in the basal ganglia. The cortico-striato-thalamo-cortical loops (CSTC loops) responsible for selection of movements are just the best known of many CSTC loops which connects the striatum with many different areas in the neocortex17. Less is known of the function of the rest of the loops, but striatum is thought to function as a dopamine dependent filter across many different modalities.
OCD and corticostriatal circuits
CSTC loops (variously involving the thalamus or the subthalamic nucleus) are heavily implicated in several theories of obsessive compulsive disorder (OCD), the evidence supporting this comes from functional neuroimaging in man as well as mice. Optogenetic studies in mice have established a causative relationship between modulation of orbitofrontal corticostriatal circuits and OCD-like symptoms (obsessive grooming behavior). OCD symptoms are affected by antipsychotics and several PET-studies of OCD in man show signs of dysregulated dopamine balance in striatum. Furthermore, extreme OCD can be treated by deep brain stimulation where the electrodes are implanted into frontorstriatal targets and also by psychosurgery where the internal capsule (a white matter tract connecting the basal ganglia and prefrontal cortex) is lesioned with directed gamma radiation 18,19.
What is OCD? OCD is a strong compulsion to do repetitive unreasonably behavior over and over again. It can also be a purely mental repetitive thoughts such as praying or counting. OCD can even include intrusive thoughts.18 What are delusions? Delusions are strong, compelling unreasonable thoughts which churn away inside the mind over and over again, always present in the background, threatening to overwhelm the afflicted20. It's plausible that if OCD emerges due to run-away feedback in CSTC loops delusions would also be able to emerge due to run-away feedback in different CSTC loops.
Delusions and the dorsal striatum
I propose that the striatum acts as a filter for selecting reasonable abstract associations and that a hyperdopaminergic tone here leads to unreasonable abstract associations. One example of an unreasonable association could be the afflicted believing that the reptile men controls his thoughts through radio waves transmitted by his dental implants. This leaky filter in striatum would be the neural correlate of delusional mood, it would explain why the afflicted fails to form logically coherent and plausible associations of the abstract relationships governing society and the wider world. However, it does not explain the iron-hard, all-consuming strength of delusions in most schizophrenics nor the rigid delusions which can occur during a trip. Therefore I also propose a related mechanism to explain the all-consuming strength of long lasting delusions. I propose that for a delusion to turn rigid a new CSTC loop with abnormally strong and consistent activity must be formed, first with Short Term Potentiation (STD) but then with long term potentiation (LTP) and in the end through more permanent neuroplasticity (new synapses, dendrites and neurites formed). The constant activity in this loop would be enabled due to the neurochemical conditions of the psychedelic brain. To be specific plasticity in the neocortex induced by 5-HT2A stimulation combined by a hyperdopaminergic signaling in the dorsal striatum. The formation of the loop would also be enabled by disinhibition from hierarchically higher areas (probably the dorsolateral prefrontal cortex). See Figure 1. An explanation for how this disinhibition occurs follows in the section on REBUS bellow. However in the cases where the delusion persists after the hallucinogen has been metabolized this would be because the neuroplasticity has been so strong that a tonic neural activity has formed in the circuit even without a global hyperserotonergic and hyperdopaminergic state. Much like tinnitus is thought to be the result of tonic activity in auditory areas due to aberrant neuroplasticity induced by the death of hair cells in the cochlea.
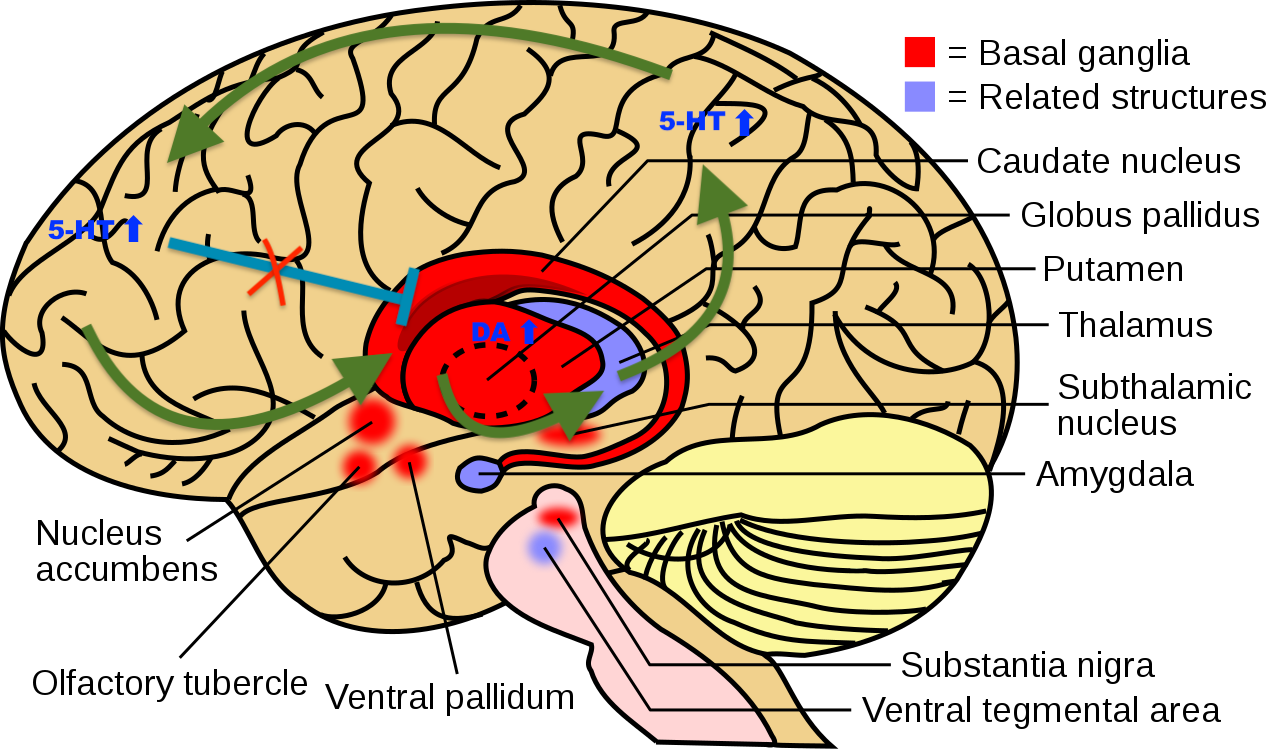
Figure 1: A simplified schematic of the hypothesized selfreinforcing CSTC-feedback-loop responsible for psychedelic-induced delusions. In this example the loop goes from the frontal lobe, to the dorsal striatum, to the thalamus, to the parietal lobe and then back to the frontal lobe. The loop becomes self reinforcing due to neuroplasticity induced by serotonergic signalling in the neocortex coupled with a hyperdopaminergic tone in striatum. The serotonergic signalling in the neocortex leads to increased neuroplasticity and disturbs cortical inhibition of striatum.
Image adapted from Wikipedia: BrainCaudatePutamen.svg: User:LeevanjacksonDerivative work: User:SUM1 - Derivative work based on File:BrainCaudatePutamen.svg, CC BY-SA 4.0, https://commons.wikimedia.org/w/index.php?curid=85845448
If you could find and sever the CSTC loop, you could sever the delusion. However, the aberrant loop is likely right next to loops encoding several useful associations, so such precision neural microsurgery is unlikely in the near future. On the other hand, if you can justify lesioning CSTC loops with a gamma knife in severe OCD, then you should be able to justify the same procedure on different loops as a treatment for severe delusions. Probably not for hallucinogen-induced delusions but definitely for some severe forms of schizophrenia.
There’s a lot of evidence supporting a crucial role of the dorsal striatum in delusions and attentional salience. Anatomically the dorsal striatum consists of caudate and putamen.20 Here CSTC loops projects from areas in the frontal lobe (in particular the dorsolateral prefrontal cortex via the dorsal striatum to areas in the parietal lobe) 20–25. Tractographic studies in humans show that the dorsal striatum functions as an integrative hub, since it receives input from many different areas in the cortex and the rest of striatum. Loops passing through the dorsal striatum from the prefrontal cortex are essential for associative learning, complex social cognition and language21. Dysfunction in these domains seems essential for the formation of most human delusions. Furthermore, people with psychosis and those at risk for psychosis have the highest increase of dopamine in this region of striatum as measured with PET-studies using radio-labelled Levodopa as a challenge. fMRI studies have also correlated specifically greater activity in the dorsal striatum to psychotic symptoms20. Therefore, it seems reasonable to start looking for the neural correlates of delusions in CSTC loops passing through dorsal striatum. Which areas of the neocortex the aberrant loops passes through seems less clear but a good starting point would be to look for it in areas in the frontal lobe and the parietal lobe.
How REBUS and the anarchic brain sets the stage for self-reinforcing CSTC loops
One hypothesis of the general action of psychedelics on the psyche which has gathered a lot of support is REBUS (Relaxed Beliefs Under pSychedelics). In short the hypothesis states that psychedelics induce a state where the output from bottom up sensory areas and the limbic system overwhelms hierarchical top down control. The top down inhibition and top down attempts to minimize prediction errors gets drastically reduced as hierarchically higher areas (such as the default mode network) gets desynchronized. There’s some limited brain imaging evidence for this but most evidence supporting the hypothesis comes from phenomenology. Psychedelics can break down established belief structures, ideological frameworks and other preconceived notions of the world. Furthermore according to the entropic brain hypothesis, psychedelics increases the electrochemical entropy in the brain which leads it closer to criticality. Criticality being defined as a neurophysiological condition where more brain states can be reached for less energy and cascade effects can avalanche through the system26,27. The phenomenological correlate for this would be that it takes less effort to think new thoughts, or that mood can swing on a dime and that abstract reasoning can spiral away quickly. The entropic brain hypothesis complements REBUS fairly well.
However the full range of psychedelic experiences is more than just relaxed beliefs and relaxed preconceived notions. In one moment one can be blissfully harmonic, having a high degree of self-dissolution, in another one can be certain one has reached the pinnacle of enlightenment and figured it all out (delusional mood or what Ann Shulgin would call Inflation28), only for a slight disturbance to send one into a downward spiral into increasingly paranoid and malicious delusions which can be very hard to break. People under the influence can spend hours (which for them can be years in subjective time) in nasty thought loops or on rigid delusions such as:
“I’m the turnip” or “Everyone around me are shrimps”. One of Huxley’s essays on his experience with mescaline wast titled “Heaven and hell” for a reason. So psychedelics can lead to criticality when the conditions are right, but when the electrochemical cascade effects spiral out of control they can also lead to very rigid thought structures which leaves long lasting imprints in the idea world of the user. This rigidity of thought stands in stark contrast to REBUS and it’s not dealt with in its entirety by the articles written by Carhart Harris et al. on the entropic brain hypothesis.
I believe that the mental instability perceived whilst under the influence is due to reduced top down control of mental homeostasis, this in turn would be aligned with the principle of REBUS. Several hierarchically higher order networks are overwhelmed by bottom up signalling, possibly due to a psychedelics-induced malfunctioning filter function in the thalamus, and are thus unable to fulfil their function of regulating prediction error, such as in delivering feedback to the dorsal striatum on what associations are reasonable. As mentioned above, the delusional mood most likely occur due to the secondary dopaminergic effects of psychedelics in striatum, and it’s likely further facilitated by lack of top down control from other areas (likely areas in the frontal lobe, particularly the dorsolateral prefrontal cortex). However, for the delusions to crystallize into something rigid which lasts for hours and defies all attempts at reason something else must happen. I believe that the combination of 5-HT2A induced neural plasticity in cortical areas and hyperdopaminergic signalling in striatum would be the driving force for this. It would be this interplay of dopaminergic and serotonergic signalling which enables the loop to become hyperactive. See Figure 1.
The amygdala and paranoia
The amygdala is so neurally interconnected with the rest of the striatum that it’s considered part of it. People with lesions to the amygdala display fearless risk taking behavior and this brain area has been strongly associated with fear and threat response in neuroscience textbooks. Due to these facts I originally hypothesized that the paranoia of paranoid delusions would be a result of a dopamine driven self-reinforcing feedback loop between the amygdala and the rest of striatum, which would integrate with the CSTC loops responsible for delusions. In this manner fear itself would integrate into the delusions making them more paranoid and sinister causing more fear (more activity in the amygdala) which would result in an even more paranoid and sinister quality to the delusions. This would quite neatly explain the negative paranoid spiral so characteristic of bad trips and endogenous paranoid psychosis. It would make intuitive sense that psychedelics would give rise to positive feedback loops across multiple different modalities in multiple different locations of the brain, feedback loops in circuits which resemble each other in physiology and receptor composition but computes mental functions in different domains of the psyche. In fact my hypothesis for the neural correlates of delusions was born out another hypothesis for complex hallucinations:
That complex serotonin driven hallucinations arise due to self-reinforcing positive feedback loops between higher and lower areas of the ventral stream of visual processing. More of that idea to be disclosed in a future article I think.
However, cutting edge neuroscience is more often than not characterized by noisy data and complex counter-intuitive relationships. Most but not all neuroimaging studies comparing paranoid to non-paranoid persons with schizophrenia show lower activity of the amygdala in the paranoid group29–33. And indeed activity in the basolateral amygdala appears to lower dopamine flow in parts of striatum by dampening the activity of dopaminergic neurons in the ventral tegmental area34. While there is plenty of neuroimaging evidence for an altered functional connectivity of the amygdala in schizophrenia35–38, the alterations appears contradictory and defies reductionist explanations. The heterogeneous results in the literature on altered amygdala connectivity doesn’t seem to be due to the noise fishing so prominent in the neuroimaging fields, rather it seems that the function of the amygdala in health and psychopathology is far more complex than the textbook definition of what an amygdala does. While it seems likely that the amygdala is involved somehow, the exact role that this brain area has to play in paranoid delusions is not yet known. I haven’t encountered any convincing theories in my research. However dopaminergic activity in the dorsal striatum has been correlated to threat perception in rats39. This may indicate that paranoia can also arise due to inherent activity in the the dorsal striatum20.
The relevance of CSTC loops during psychedelic therapy
The current doctrine in psychedelic therapy is for the therapist to sit through a psychedelic session, not to actively steer it. While it’s sensible to not disturb the patient while he is quietly working through past trauma or experiencing self-dissolution, it seems unwise to leave an emerging delusion/self-reinforcing-CSTC loop untreated. If my hypothesis is correct, the longer it gets to go on the more rigid the aberrant association will become. This would be due to STP turning into LTP (upregulation of enzymes producing neurotransmitters and neurotrophins) turning into construction of new dendrites, axons and synapses. Though more research is needed on the frequency of persistent delusions as a result of hallucinogen use, in theory it would be best to break the loop while it’s still in the early looser stage, before it gets tonically active. Of course the best of all would be to prevent paranoid delusions from forming in the first place through correct set and setting, space holding, choice of music etc. However, therapy is a craft not a science, and psychedelic therapy is an emerging and experimental craft. Mistakes will be made. If the therapist notices a malicious delusion which is persistent over time there should be a chance to break the correlating loop through distraction, a rapid change in mindset or physical setting will change the electrochemical environment in the brain, this in turn may break the loop.
The impact of dysfunctional dopaminergic signalling on psychedelic culture
Long term psychotic symptoms, (lasting for days or months) can occur in conjunction with clandestine use of classical psychedelics. It’s highly overrepresented among hallucinogen users who suffer from, or have a family history of schizophrenic and/or bipolar disorders. However, sometimes long term psychotic symptoms emerge even in users who don't even have a family history of the disorders mentioned above40–43. Large online survey studies and meta analysis of available literature shows that the incidence of long term psychosis in conjunction with use is exceedingly rare even in clandestine settings, but does indeed occur sometimes42,44. Carbonaro et. al included 1339 persons into a large online survey because they’d reported challenging experiences during the use of psychedelics. When queried on their single worst psychedelic experience 9% reported having paranoia which lasted for more than one week after the trip42. Due to the nature of the surveys and meta-analyses and the unknown variables associated with retrospective research into illegal activities, a causative link between psychedelics and psychotic symptoms couldn’t be definitely proven. Indeed I can’t think of a way that a causative link can be definitely proven as long as these drugs are illegal, you just can’t get a clean sample size properly controlled for all the confounding factors. However, by reviewing the evidence it seems highly likely to me that psychedelics can catalyze primarily symptoms such as paranoia and derealization, but also more infrequently psychotic episodes. I would ask anyone with a different view this:
What more evidence (which can be gathered under current juridical conditions) do you need to see to believe in a causative link?
It’s also worth noting that the really paranoid individuals would be disinclined to answer online surveys about their drug use and paranoia because they’d think the survey was a spy-tool used by whatever hostile force they imagine are out to get them. They’d also be disinclined to seek healthcare for the same reason. One of the two large surveys I studied in my research was also advertised mostly on psychedelic-inclined internet forums which obviously would result in recruiting mostly people with a positive attitude towards these drugs42.
I propose that in the few cases where a bad trip flowers out into fullblown delusions lasting well after the particular drug has been metabolized it’s because aberrant CSTC loops, passing through the dorsal striatum (representing aberrant delusions), have formed and become tonically active. Neurons who fire together wire together and the psychedelic’s stimulation of 5-HT2A in the neocortex along with hyperdopaminergic signalling in striatum during the trip have caused neurons who shouldn’t fire together to wire together and form new synapses. For the less rigid aberrant beliefs and personality changes in the user community, perhaps the new aberrant circuits are not tonally active but hypersensitized, ready to fire at the slightest internal or external trigger. Rather than an all or nothing concept, consider the neuroplastic changes in the CSTC loops to be a full spectrum, from slight sensitization to tonal activity. This would correspond to the full spectrum of change in the idea-world of users, from full blown delusions via flashbacks to slightly aberrant ideas which are stronger than usual.
This would go a long way to explain the tendency in psychedelic culture towards paranoid conspiracy theories and these CSTC loops together with the memory of dopamine-driven delusional mood would go a long way to explain the development of intense faith in magical belief structures, such as those prevalent in the new age movement. For the sake of intellectual honesty it should be noted that the people who use hallucinogens are also often using cannabis and amphetamines, compounds who are known to cause paranoia and delusions for a significant part of their users45–47.
What are the long term complications of having a transient period of attentional salience and delusional mood? One hypothesis is that this, combined with the immediacy of having one’s default mode network desynchronized and a bottom up overflow of sensory perception, is that the world, as perceived after the hallucinogen has left the body, might pale in comparison to the trip-world. It would not surprise me if dopamine had a role to play in derealization, which is an unwanted side effect which sometimes occurs after the use of psychedelics. In a small interview study which specifically searched for individuals who had experienced long term psychological harm which they attributed to their use of classical psychedelics, derealization was present in 7 out of 15 participants for well over 72 hours after the trip48. A part of this derealization is probably due to poorly integrated self dissolution, but a part of this is also likely due to clear memories of the world as perceived during delusional mood and the vividness of the colors and shapes (a memory of dopaminergic attentional salience in combination with serotonergic sensory overflow). Derealization is a poor term to describe it really, lack of hyperrealization would be more appropriate. This could be an explanation for why so many people believe that the complex hallucinations perceived during a DMT-trip don’t originate from the brain but rather are a view into some kind of parallel universe, or that they’re communicating with aliens. The trip feels realer than reality so people jump to conclusions.
One could also imagine that delusional mood and attentional salience together with neuroplasticity and loss of top down inhibition could cause the user to ascribe an unreasonable amount of significance to certain objects, persons and ideas who are present during the psychedelic experience. This could lead to suboptimal priorities in life. If psychedelics can cause stable, tonally active and abnormal CSTC-loops which correlate to delusions, then MKUltra was probably (to some extent) correct in their idea that LSD can be used as a brain washing agent. It is worth noting that several cults have used psychedelics to break down their follower’s belief structures, idea-worlds and sense of self in order to imprint unto them the values and idea-world of the cult. Aum Shrinyoku, the Manson Family, The Family (led by Anne Hamilton-Byrne) to mention a few. The spiritual narcissism which is present in psychedelic (cult)ure is also likely directly attributable to induced neuroplasticity in combination with dopamine overflow. Delusions of grandeur are often present in manic psychosis and are strongly correlated to excess dopaminergic tone. This would correlate well what Ann Shulgin described as inflation:
A state of imagined clarity where the user believes she has figured out all the secrets of the universe.28 Another example of inflation would be Dennis McKenna's oracle trip in “True Hallucinations”49.
Psychedelic therapy and psychedelic cults
Unchecked spiritual narcissism, particularly in charismatic individuals, could become quite toxic in a society as void of spirituality as the West. When the psilocybin-clinics for the severely depressed opens up, I suspect a new kind of pharmacovigilance will be needed. A pharmacovigilance to prevent hypomanic or derealized former psilocybin patients to become ample prey from psychedelic fuelled cults in their quest for meaning in life. I suspect they’re already a risk group for joining cults since cults provide a strong sense of meaning and lack of perceived meaning in life are one of the most challenging aspects of depression. There’s also an association between depression and insecure attachment styles. Spiritual awakening in people with insecure attachment styles (as compared to people with secure attachment styles) more often leads to a fervent yet unstable religious conversion with the divine as an unreliable attachment object, or in the case of people with the insecure attachment style categorized as disorganized/unresolved, spiritual awakening comes with a greater tendency towards dissociative mystical experiences.50 I’m concerned that these groups would be particularly vulnerable to recruitment attempts from future psychedelically fuelled cults.
It’s worth noting that delusional mood remains a much sought after mystical state by many religious practitioners across the globe, and mystical experiences have been correlated by many studies to the therapeutic benefit of psychedelics. A psilocybin-induced spiritual awakening is not something that can or should be put back into the subconscious box and forgotten about. When a spiritual awakening occurs as a result of psychedelic therapy and it becomes obvious that the patient wants to become part of a larger religious or mystic community, it would be a wise harm-reduction policy to present a bufé to patient of religious or spiritual congregations which are deemed harmless to society and the patient himself. It would be most appropriate to do this in conjunction with therapy, either by a non-profit foundation dedicated to the task, or by the care-provider itself. Remember, psychedelic therapy is politically controversial, there are significant conservative elements of society which would like to see both psychedelic therapy and psychedelic research shut down. We get one drug fuelled Jonestown, possibly two, and then there’ll be crackdowns and no more hallucinogen research for anyone.
Suggestions for future research
How would we go about discovering CSTC loops generating delusions? While rat models might do for OCD at a stretch I sincerely doubt rats, with their smooth neocortices, are capable of the abstract reasoning to come up with that the cultural elite are smoking DMT on the moon in order to pave the way for interdimensional soul-sucking vampires. You could ask them about it but they wouldn’t respond. For that reason I’m highly sceptical of using animals as a model system for delusions. EEG has an appropriate time resolution for tracking the progression of the action potential around the loop but its spatial resolution is abysmal. In addition to this it would take machine learning to look at anything below the neocortex and those analysis methods are quite experimental yet. It’s possible the loops can be found in humans using diffusion tensor imaging (DTI) combined with (sufficiently statistically powered and correct) fMRI. Receptor occupancy challenges measured by PET and using radiolabeled challengers such as levodopa, selective dopamine antagonists and selective serotonin agonists would be the main method for measuring the aberrant neurochemistry in vivo. If this was combined with transcranial magnetic stimulation (TMS) over the relevant area in the cortex in order to desynchronize neural firing it’s possible that the loop can be diminished. There’s an unknown risk that this might cause an epileptic seizure. Ethical approval will probably not be granted. Once the loop establishes itself in the brain it might be refined to a very slim yet very hyperactive circuit. It might be as narrow as ten axons wide at its narrowest extent. Therefore there’s a certain risk that the loops can’t be measured with current non-invasive methods.
An interesting tangent to the research that went into producing this text is that severe delusions can probably be treated by psychosurgery. To be more specific, deep brain stimulation (DBS) in the dorsal striatum in order to disrupt the aberrant CSTC loops. Of course invasive neurosurgery always brings with it a risk of bacterial infection of the brain, but seeing as how around 10% of people suffering from schizophrenia commit suicide anyway and 10-30% don’t respond to antypsychotics25 I believe the risk is warranted. I wish I was the first to come up with this but alas, DBS in other brain areas has already been used in experimental settings as a treatment for treatment resistant schizophrenia and the dorsal striatum has been pointed out as a good target before.25 However, to my knowledge no one has actually gone through with the surgery and used DBS at this brain area to treat either schizophrenia or delusions yet. It would probably zombify people less than antipsychotics since the former would be a less systemic treatment. Of course, proper PET/DTI evaluation needs to be conducted before surgery in order to be reasonably certain of disturbed activity in the dorsal striatum in the relevant individuals. After all, no two schizophrenics are alike.
I welcome all informed academic discourse
References
1. Watts, V.J., Mailman, R.B., Lawler, C.P., Neve, K.A., and Nichols, D.E. (1995). LSD and structural analogs: pharmacological evaluation at D1 dopamine receptors. Psychopharmacology (Berl.) 118, 401–409. 10.1007/BF02245940.
2. Minuzzi, L., Nomikos, G.G., Wade, M.R., Jensen, S.B., Olsen, A.K., and Cumming, P. (2005). Interaction between LSD and dopamine D2/3 binding sites in pig brain. Synap. N. Y. N 56, 198–204. 10.1002/syn.20141.
3. Nichols, D.E. (2016). Psychedelics. Pharmacol. Rev. 68, 264–355. 10.1124/pr.115.011478.
4. Ray, T.S. (2010). Psychedelics and the Human Receptorome. PLOS ONE 5, e9019. 10.1371/journal.pone.0009019.
5. Howell, L.L., and Cunningham, K.A. (2015). Serotonin 5-HT2 Receptor Interactions with Dopamine Function: Implications for Therapeutics in Cocaine Use Disorder. Pharmacol. Rev. 67, 176–197. 10.1124/pr.114.009514.
6. Inserra, A., De Gregorio, D., and Gobbi, G. (2021). Psychedelics in Psychiatry: Neuroplastic, Immunomodulatory, and Neurotransmitter Mechanisms. Pharmacol. Rev. 73, 202–277. 10.1124/pharmrev.120.000056.
7. Vollenweider, F.X., Vontobel, P., Hell, D., and Leenders, K.L. (1999). 5-HT modulation of dopamine release in basal ganglia in psilocybin-induced psychosis in man--a PET study with [11C]raclopride. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 20, 424–433. 10.1016/S0893-133X(98)00108-0.
8. Albizu, L., Holloway, T., González-Maeso, J., and Sealfon, S.C. (2011). Functional crosstalk and heteromerization of serotonin 5-HT2A and dopamine D2 receptors. Neuropharmacology 61, 770–777. 10.1016/j.neuropharm.2011.05.023.
9. Borroto-Escuela, D.O., Romero-Fernandez, W., Narvaez, M., Oflijan, J., Agnati, L.F., and Fuxe, K. (2014). Hallucinogenic 5-HT2AR agonists LSD and DOI enhance dopamine D2R protomer recognition and signaling of D2-5-HT2A heteroreceptor complexes. Biochem. Biophys. Res. Commun. 443, 278–284. 10.1016/j.bbrc.2013.11.104.
10. Guevara-Lora, I., Niewiarowska-Sendo, A., Polit, A., and Kozik, A. (2016). Hypothetical orchestrated cooperation between dopaminergic and kinin receptors for the regulation of common functions. Acta Biochim. Pol. 63, 387–396. 10.18388/abp.2016_1366.
11. Łukasiewicz, S., Błasiak, E., Szafran-Pilch, K., and Dziedzicka-Wasylewska, M. (2016). Dopamine D2 and serotonin 5-HT1A receptor interaction in the context of the effects of antipsychotics – in vitro studies. J. Neurochem. 137, 549–560. 10.1111/jnc.13582.
12. Mishara, A.L., and Fusar-Poli, P. (2013). The Phenomenology and Neurobiology of Delusion Formation During Psychosis Onset: Jaspers, Truman Symptoms, and Aberrant Salience. Schizophr. Bull. 39, 278–286. 10.1093/schbul/sbs155.
13. Bell, V., Raihani, N., and Wilkinson, S. (2021). Derationalizing Delusions. Clin. Psychol. Sci. 9, 24–37. 10.1177/2167702620951553.
14. Stegmayer, K., Horn, H., Federspiel, A., Razavi, N., Bracht, T., Laimböck, K., Strik, W., Dierks, T., Wiest, R., Müller, T.J., et al. (2013). Ventral striatum gray matter density reduction in patients with schizophrenia and psychotic emotional dysregulation. NeuroImage Clin. 4, 232–239. 10.1016/j.nicl.2013.12.007.
15. Hilty, D.M., Leamon, M.H., Lim, R.F., Kelly, R.H., and Hales, R.E. (2006). A Review of Bipolar Disorder in Adults. Psychiatry Edgmont 3, 43–55.
16. Bamford, N.S., Zhang, H., Schmitz, Y., Wu, N.-P., Cepeda, C., Levine, M.S., Schmauss, C., Zakharenko, S.S., Zablow, L., and Sulzer, D. (2004). Heterosynaptic Dopamine Neurotransmission Selects Sets of Corticostriatal Terminals. Neuron 42, 653–663. 10.1016/S0896-6273(04)00265-X.
17. Obeso, J.A., Rodríguez-Oroz, M.C., Benitez-Temino, B., Blesa, F.J., Guridi, J., Marin, C., and Rodriguez, M. (2008). Functional organization of the basal ganglia: Therapeutic implications for Parkinson’s disease. Mov. Disord. 23, S548–S559. 10.1002/mds.22062.
18. Barahona-Corrêa, J.B., Camacho, M., Castro-Rodrigues, P., Costa, R., and Oliveira-Maia, A.J. (2015). From Thought to Action: How the Interplay Between Neuroscience and Phenomenology Changed Our Understanding of Obsessive-Compulsive Disorder. Front. Psychol. 6, 1798. 10.3389/fpsyg.2015.01798.
19. Balachander, S., Arumugham, S.S., and Srinivas, D. (2019). Ablative neurosurgery and deep brain stimulation for obsessive-compulsive disorder. Indian J. Psychiatry 61, S77–S84. 10.4103/psychiatry.IndianJPsychiatry_523_18.
20. McCutcheon, R.A., Abi-Dargham, A., and Howes, O.D. (2019). Schizophrenia, Dopamine and the Striatum: From Biology to Symptoms. Trends Neurosci. 42, 205–220. 10.1016/j.tins.2018.12.004.
21. Sabaroedin, K., Tiego, J., and Fornito, A. (2023). Circuit-Based Approaches to Understanding Corticostriatothalamic Dysfunction Across the Psychosis Continuum. Biol. Psychiatry 93, 113–124. 10.1016/j.biopsych.2022.07.017.
22. Gremel, C.M., and Lovinger, D.M. (2017). Associative and Sensorimotor Cortico-Basal Ganglia Circuit Roles in Effects of Abused Drugs. Genes Brain Behav. 16, 71–85. 10.1111/gbb.12309.
23. Dandash, O., Pantelis, C., and Fornito, A. (2017). Dopamine, fronto-striato-thalamic circuits and risk for psychosis. Schizophr. Res. 180, 48–57. 10.1016/j.schres.2016.08.020.
24. Levitt, J.J., Nestor, P.G., Levin, L., Pelavin, P., Lin, P., Kubicki, M., McCarley, R.W., Shenton, M.E., and Rathi, Y. (2017). Reduced Structural Connectivity in Frontostriatal White Matter Tracts in the Associative Loop in Schizophrenia. Am. J. Psychiatry 174, 1102–1111. 10.1176/appi.ajp.2017.16091046.
25. Mikell, C.B., Sinha, S., and Sheth, S.A. (2016). Neurosurgery for schizophrenia: an update on pathophysiology and a novel therapeutic target. J. Neurosurg. 124, 917–928. 10.3171/2015.4.JNS15120.
26. Carhart-Harris, R.L., Leech, R., Hellyer, P.J., Shanahan, M., Feilding, A., Tagliazucchi, E., Chialvo, D.R., and Nutt, D. (2014). The entropic brain: a theory of conscious states informed by neuroimaging research with psychedelic drugs. Front. Hum. Neurosci. 8. 10.3389/fnhum.2014.00020.
27. Carhart-Harris, R.L. (2018). The entropic brain - revisited. Neuropharmacology. 10.1016/j.neuropharm.2018.03.010.
28. Shulgin A. and Shulgin A. (1997). Tihkal: The Continuation, Chapter 10, page 165, ISBN 0-9630096-9-9
29. Pinkham, A.E., Liu, P., Lu, H., Kriegsman, M., Simpson, C., and Tamminga, C. (2015). Amygdala Hyperactivity at Rest in Paranoid Individuals With Schizophrenia. Am. J. Psychiatry 172, 784–792. 10.1176/appi.ajp.2014.14081000.
30. Williams, L. (Lea) M., Das, P., Liddell, B.J., Olivieri, G., Peduto, A.S., David, A.S., Gordon, E., and Harris, A.W.F. (2007). Fronto-limbic and autonomic disjunctions to negative emotion distinguish schizophrenia subtypes. Psychiatry Res. Neuroimaging 155, 29–44. 10.1016/j.pscychresns.2006.12.018.
31. Williams, L.M., Das, P., Harris, A.W.F., Liddell, B.B., Brammer, M.J., Olivieri, G., Skerrett, D., Phillips, M.L., David, A.S., Peduto, A., et al. (2004). Dysregulation of arousal and amygdala-prefrontal systems in paranoid schizophrenia. Am. J. Psychiatry 161, 480–489. 10.1176/appi.ajp.161.3.480.
32. Russell, T.A., Reynaud, E., Kucharska-Pietura, K., Ecker, C., Benson, P.J., Zelaya, F., Giampietro, V., Brammer, M., David, A., and Phillips, M.L. (2007). Neural responses to dynamic expressions of fear in schizophrenia. Neuropsychologia 45, 107–123. 10.1016/j.neuropsychologia.2006.04.026.
33. Phillips, M.L., Williams, L., Senior, C., Bullmore, E.T., Brammer, M.J., Andrew, C., Williams, S.C., and David, A.S. (1999). A differential neural response to threatening and non-threatening negative facial expressions in paranoid and non-paranoid schizophrenics. Psychiatry Res. 92, 11–31. 10.1016/s0925-4927(99)00031-1.
34. Sonnenschein, S.F., Gomes, F.V., and Grace, A.A. (2020). Dysregulation of Midbrain Dopamine System and the Pathophysiology of Schizophrenia. Front. Psychiatry 11, 613. 10.3389/fpsyt.2020.00613.
35. Liu, H., Tang, Y., Womer, F., Fan, G., Lu, T., Driesen, N., Ren, L., Wang, Y., He, Y., Blumberg, H.P., et al. (2014). Differentiating Patterns of Amygdala-Frontal Functional Connectivity in Schizophrenia and Bipolar Disorder. Schizophr. Bull. 40, 469–477. 10.1093/schbul/sbt044.
36. Tian, L., Meng, C., Yan, H., Zhao, Q., Liu, Q., Yan, J., Han, Y., Yuan, H., Wang, L., Yue, W., et al. (2011). Convergent Evidence from Multimodal Imaging Reveals Amygdala Abnormalities in Schizophrenic Patients and Their First-Degree Relatives. PLOS ONE 6, e28794. 10.1371/journal.pone.0028794.
37. Kim, W.-S., Shen, G., Liu, C., Kang, N.-I., Lee, K.-H., Sui, J., and Chung, Y.-C. (2020). Altered amygdala-based functional connectivity in individuals with attenuated psychosis syndrome and first-episode schizophrenia. Sci. Rep. 10, 17711. 10.1038/s41598-020-74771-w.
38. Jalbrzikowski, M., Murty, V.P., Tervo-Clemmens, B., Foran, W., and Luna, B. (2019). Age-Associated Deviations of Amygdala Functional Connectivity in Youths With Psychosis Spectrum Disorders: Relevance to Psychotic Symptoms. Am. J. Psychiatry 176, 196–207. 10.1176/appi.ajp.2018.18040443.
39. Menegas, W., Akiti, K., Amo, R., Uchida, N., and Watabe-Uchida, M. (2018). Dopamine neurons projecting to the posterior striatum reinforce avoidance of threatening stimuli. Nat. Neurosci. 21, 1421–1430. 10.1038/s41593-018-0222-1.
40. Barber, G., Nemeroff, C.B., and Siegel, S. (2022). A Case of Prolonged Mania, Psychosis, and Severe Depression After Psilocybin Use: Implications of Increased Psychedelic Drug Availability. Am. J. Psychiatry 179, 892–896. 10.1176/appi.ajp.22010073.
41. Dos Santos, R.G., Bouso, J.C., and Hallak, J.E.C. (2017). Ayahuasca, dimethyltryptamine, and psychosis: a systematic review of human studies. Ther. Adv. Psychopharmacol. 7, 141–157. 10.1177/2045125316689030.
42. Carbonaro, T.M., Bradstreet, M.P., Barrett, F.S., MacLean, K.A., Jesse, R., Johnson, M.W., and Griffiths, R.R. (2016). Survey study of challenging experiences after ingesting psilocybin mushrooms: Acute and enduring positive and negative consequences. J. Psychopharmacol. Oxf. Engl. 30, 1268–1278. 10.1177/0269881116662634.
43. Hendin, H.M., and Penn, A.D. (2021). An episode of mania following self-reported ingestion of psilocybin mushrooms in a woman previously not diagnosed with bipolar disorder: A case report. Bipolar Disord. 23, 733–735. 10.1111/bdi.13095.
44. Schlag, A.K., Aday, J., Salam, I., Neill, J.C., and Nutt, D.J. (2022). Adverse effects of psychedelics: From anecdotes and misinformation to systematic science. J. Psychopharmacol. Oxf. Engl. 36, 258–272. 10.1177/02698811211069100.
45. Bramness, J.G., and Rognli, E.B. (2016). Psychosis induced by amphetamines. Curr. Opin. Psychiatry 29, 236. 10.1097/YCO.0000000000000254.
46. Sanchez-Ramos, J. (2015). Chapter Seven - Neurologic Complications of Psychomotor Stimulant Abuse. In International Review of Neurobiology The Neuropsychiatric Complications of Stimulant Abuse., P. Taba, A. Lees, and K. Sikk, eds. (Academic Press), pp. 131–160. 10.1016/bs.irn.2015.02.003.
47. Burns, J.K. (2013). Pathways from cannabis to psychosis: a review of the evidence. Front. Psychiatry 4, 128. 10.3389/fpsyt.2013.00128.
48. Bremler, R., Katati, N., Shergill, P., Erritzoe, D., and Carhart-Harris, R. (2023). Focusing on the negative: cases of long-term negative psychological responses to psychedelics. 10.31234/osf.io/yzmcj.
49. True Hallucinations https://www.goodreads.com/book/show/114867.True_Hallucinations.
50. Attachment in Religion and Spirituality: A Wider View by Pehr Granqvist | Goodreads https://www.goodreads.com/book/show/52341354-attachment-in-religion-and-spirituality?from_search=true&from_srp=true&qid=0kPrsBp0oW&rank=1.